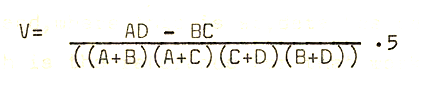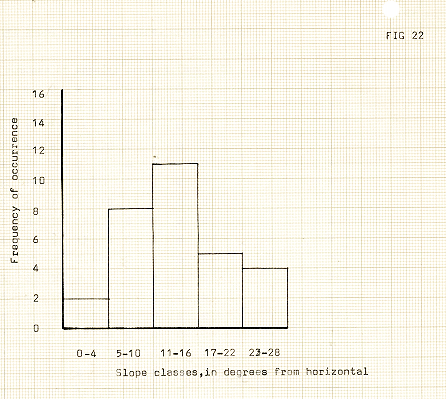'Everyone is a Ragged University - a unique and distinct body of knowledge, accredited by their life experience with a membership of one'
Plant Communities and Successional Characteristics by R . A . Cranwell
This is a digitised copy of the original research done on ‘Association Analysis of Plant Communities and Successional Characteristics on Recently Deglaciated Terrain at Svinafell, S.E. Iceland’ by Robert A. Cranwell.
R. A. Cranwell
Association Analysis of Plant Communities and Successional Characteristics on Recently Deglaciated Terrain at Svinafell, S.E. Iceland
Volume I
by R . A . Cranwell
1980
Table of Contents
Abstract
This work, based on an original study in Southeastern Iceland, identifies by the association analysis methods pioneered by Williams and Lambert, (1959), the roles and characteristics of the major plant communities inhabiting an area of recent morainic deposits. It traces the differenced in micro-environment, and in plant strategies which influence and maintain the communities.
In addition, the question of succession in this environment, previously considered by Cooper, (1916), and Tisdale, (1966), is reviewed and re-explored with specific reference to successions of Dryas spp. and Salix spp., and the relative roles of the Salix species present in the succession
Acknowledgements
Thanks go to Mr. P. Wilkinson, B.Sc., a former student of the college who helped considerably, in many different ways whilst on the Icex 79 Expedition, and without his help it is doubtful whether either of the studies would have been of their present extent without him. Thanks, too, to Mr Gordon Forrest, of the college’s computer department, for suggestions and guidance whilst using the facilities of his department.
Thanks also, to the members of the Biology/Environmental science department who, by constructive comment or merely by their interest helped in the production of this study. Finally, my thanks and the dedication of this work go to my parents, who although they may not understand this work, without their help, neither would I.
Since the beginning of this century a small number of workers in plant ecology have been impressed with the unique opportunities which recently deglaciated terrain affords for the study of Vegetational development. The earliest works, well known for their originality and scope, were those of Cooper, (1916; 1923; 1939), who carried out investigations on the moraines of the Mount Robson area of British Columbia. His later works were devoted to similar, but more extensive studies of the vegetation in the Glacier Bay sequences of Alaska. More recently, Tisdale et al., (1966), returned to the original area studied by Cooper (l916), and carried out follow-up work on the original study.
The vegetation of the area showed clear trends, of early dominance by low shrubs such as Dryas spp., and Hedysarum spp., later being succeeded in turn by Salix spp., Arctostaphylos rubra, and Betula spp. scrub. The final transitional stage was one which bore great resemblance to the surrounding forest, which was dominated by Picea engelmanni. however the composition of the vegetation differed markedly from the mature forest. Particularly of note in relation to this present study is the conclusion that phases of succession characterised by Dryas, then Salix species, were integral in the successions of deglaciated terrain of that area.
Whilst it is recognised that previous studies have been in their way invaluable for the insight they provided into succession in this environment, the data base and analysis for the conclusions reached by previous workers has often been of a more descriptive than quantitative nature. Although this present work may be somewhat lacking in volume of data, the analysis to which the data has been subject has been both extensive and rigorous. In this respect at least, it is hoped, that a more reliable and detailed appreciation of the successional changes and Vegetational strategies within plant communities of this terrain may be reached.
The field work upon which this study is based mas conducted in conjunction with work on soil genesis in recently deglaciated terrain in South-East Iceland, in July, 1979, ( Cranwell, 1979). This work involved assessment of vegetation cover, amongst other things, and in the course of the work it became clear that distinct changes mere occurring in the vegetation, both in a direction at right angles to the glacier snout, and in relation to the surface topography.
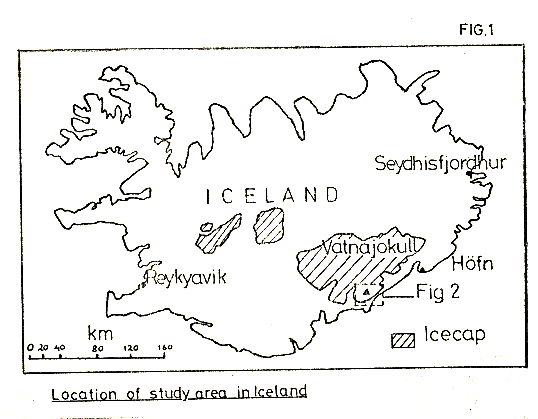
Figure 1
The latter change has very evidently the more pronounced of the two, a feature, which, in view of the remarkably hummocky nature of the terrain, mas to be expected. The extent and variation in these changes has been clarified statistically in this study, using the principles which Williams and Lambert, (1959), devised for association analysis of vegetation communities
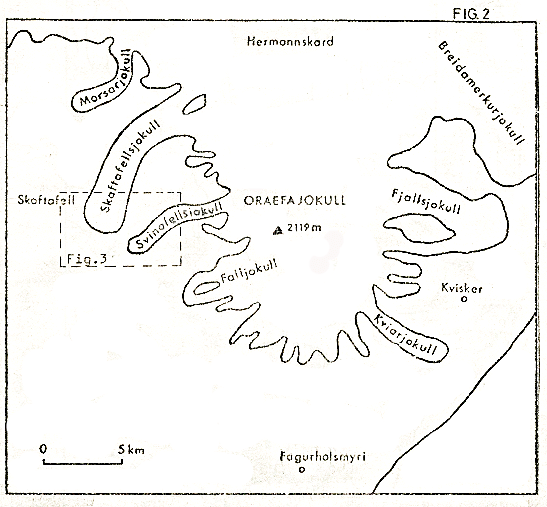
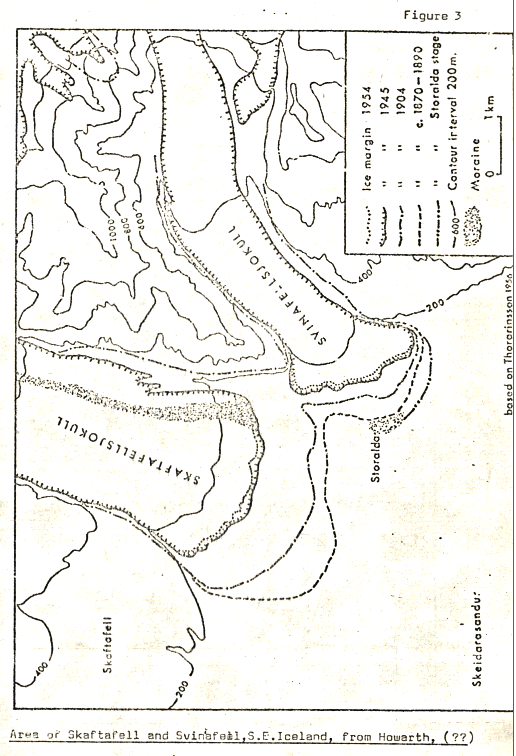
The area of study lies at the end of one of the outlet glaciers of Vatnajokull, Europe’s largest icecap, (see Figs 1, 2). The Svinafell glacier itself has experienced considerable thinning and retreat over the last eighty years of more, the snout now being some 400m further up the glacial valley than the 1904 margin. Figure 3 illustrates the extent of this recession, and, as can be seen, the glacier is bounded by sharp ridges up to 1000m in height, composed of heavily frost shattered basaltic flows.
Figure 2
At the snout, the glacier is some 150m above sea level, as are the latest moraines, however the moraines gradually decrease in height to about 100m at the start of the outwash plain itself, which then slopes very gently down to the sea, about 6-8km distant. The whole area is dominated by Hvannadalshnukur, at 2119m, Iceland’s highest peak, and part of the volcanic cone of Oraefajokull.
Figure 3
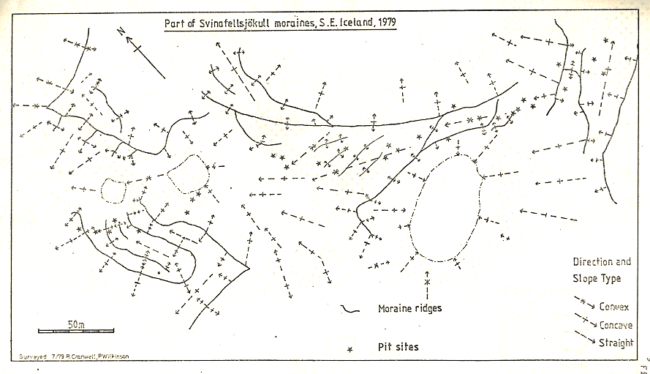
Figure 4
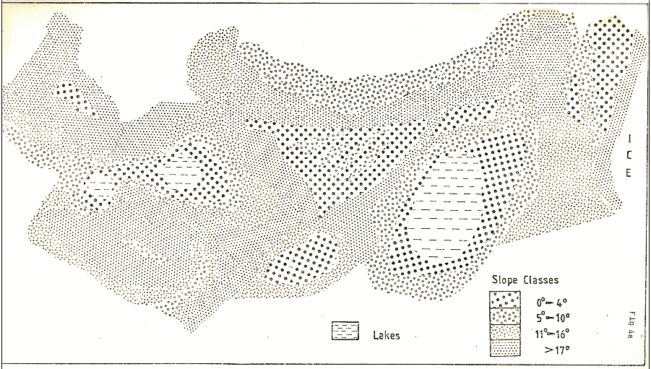
Figure 4a
FIGURE
Vegetation in the area tends to be in three main altitudinal zones:
1) above 350m, barren rock and poor alpine heath, a zone which is dominated by Rhacomitrium mosses.
2) below 350m, but above the “forest” zone are dry alpine heaths and meadows, grasses and pockets of Salix and Betula scrub. the forest zone, usually from 100 to 200 metres, predominantly Betula scrub.
The outwash plain is dominated by mosses, probably Rhacomitrium. whilst vegetation on the moraines is very patchy, determined primarily by the extremely hummocky nature of the terrain, (see fig.4, 4a).
It is the purpose of this study to try and isolate, where possible, the various communities which colonise the extremely harsh environment of the moraines, and further, to define the ways in which succession occurs in comparison to other studies in differing, but glacial areas.
Methods
After a thorough reconnaissance of the area, a transect was taken from the ice front, using what was considered to be as representative an area of the terrain as possible. The details of the area are shown in figure 4, the dimensions of the area were 400m long by 30m wide.
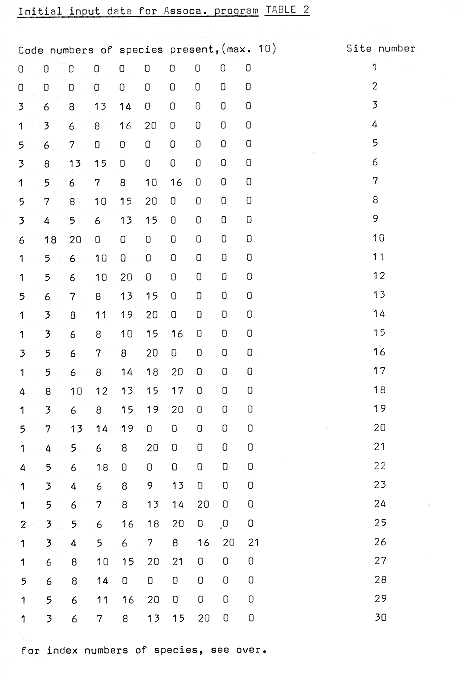
The actual sites used within the area were determined by using random number tables to identify co-ordinates for length and breadth. The method used was to divide the 400m length into 100m sections and to take numbers from the tables to identify length co-ordinates after starting from an arbitrary point within the random number table, until 30 sites were selected. To pinpoint the sites within the transect, breadth co-ordinates were then selected, using all the numbers under 30 which appeared after an arbitrary starting point, and where numbers over 30 appeared, subtracting 30, or a multiple thereof, and using the remainder as the breadth co-ordinate. The actual location of the sites was accomplished using a series of tape measures, first for location of the length co-ordinates, and then for the breadth co-ordinates.
Sampling of the vegetation was recorded using a simple percentage cover for each species present within a one metre radius of the point located by the co-ordinates. Although more sophisticated methods of recording and assessment undoubtedly exist, it was felt that the use of a familiar and practised technique would be of more immediate value than the possible confusion arising from unfamiliar techniques. In the event, the data used for the analysis was a simple presence/absence indicator, however, the original data are presented in the following section, and a number of points of interest have been noted from the percentage data.
The sample size is admittedly small, being only 30 sites, especially when the large inherent variation in the terrain is considered, however it must be noted that time was limiting, and by no stretch of the imagination could a more thorough survey have been completed in the time available, bearing in mind that extensive in situ tests were employed in the investigation of the soils of the area, and some of which information is integral in the interpretation of the Vegetational patterns of the area.
Analysis
The analysis of the data was conducted using as a basis a program devised by Messrs. Yates and Forrest of the College’s Environmental Science and Computer departments, respectively. The index of association used was the point correlation coefficient V, which is calculated as follows;
Where A = number of quadrats with both species, (of any given pair) present
B = number of quadrats with only first species present
C= number of quadrats with only second species present
D = number of quadrats with neither species present
V has a value of between +1 and -1, where 0 indicates that two species are distributed independently of one another, statistically, and +1 or -1 indicate strong positive and negative relationships respectively. The procedure follows the “normal” association analysis, devised by Williams and Lambert (1959), where the total number of quadrats is progressively broken down into discrete groups which are positively associated internally, but negatively associated, to greater or lesser degrees, externally.
After choosing the association index to be used, (V is the most widely used, (Poole; 1974; p346)), the index was then calculated for all possible pairs of species within the original data. The V-values were then summed for each species, regardless of sign, and listed on the computer printout (see Appendices). Then, if species A had the highest total V value, the data used as input for that analysis was then subdivided into two groups, on the presence or absence of species A in the quadrat. The program mas then re-run, with two new data sets as inputs, and the process mas repeated with each successive division until a hierarchical arrangement of community associations mas achieved. The end of the program run is reached when, in each sub-set of data, there are equal V-values (summed), for each of the species under consideration.
Results
The results displayed on the following pages represent in part, data which has previously been used in the dissertation for the Geography department, and, where this is so, data has been quoted as from Cranwell (1979), which is the reference for that work. In all other cases the data was acquired specifically for this dissertation and has been subject to analysis only in respect of this dissertation.
Ram data is displayed, but for ease of interpretation, the bulk of the data is presented in graphical form, and usually in conjunction with another variable, e .g., species cover against distance downslope.
Figure 5
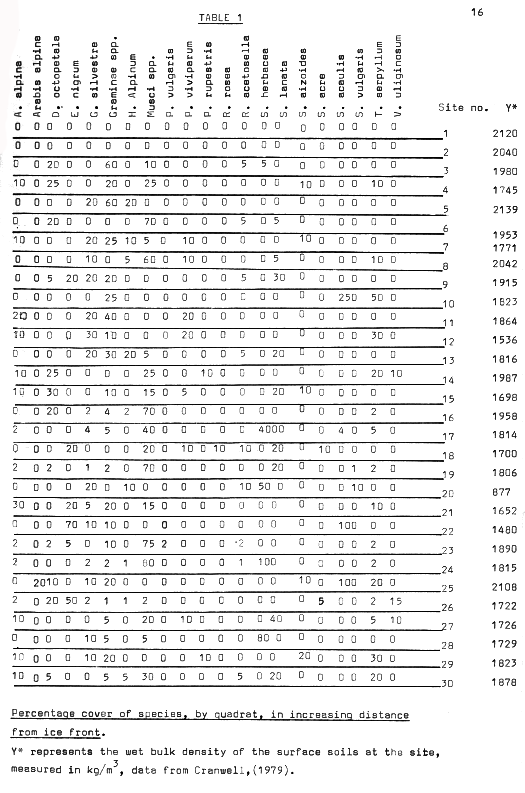
Table 1
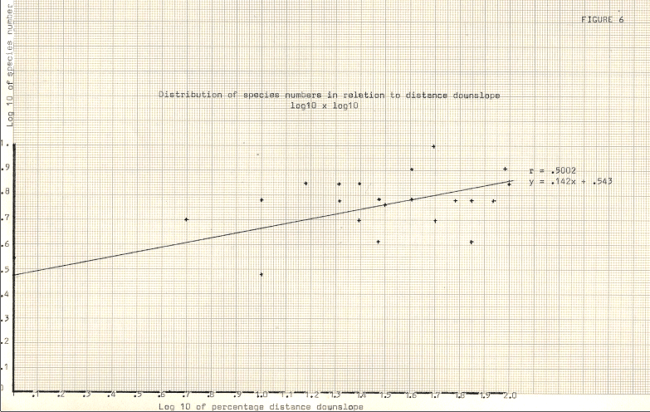
Figure 6
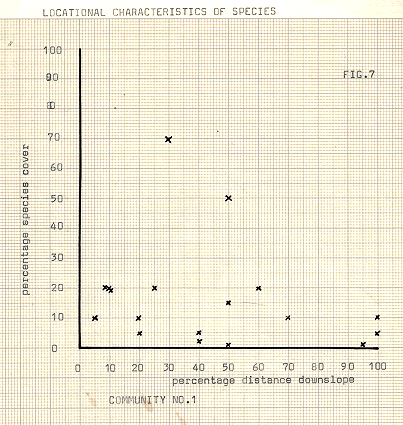
Figure 7
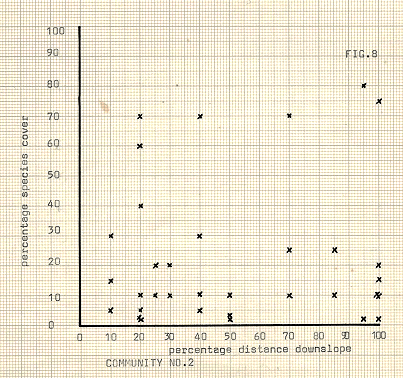
Figure 8
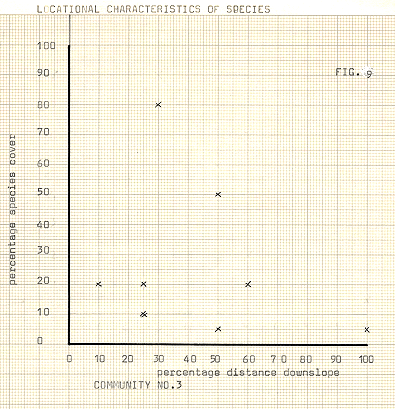
Figure 9
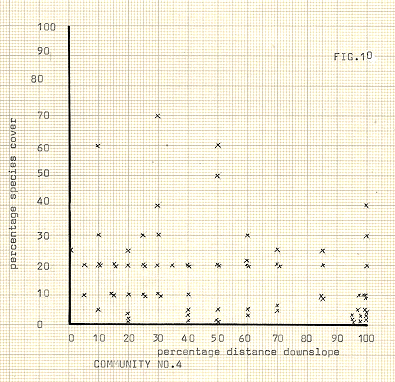
Figure 10
Figure 11
Figure 12
Figure 13
Figure 14
Figure 15
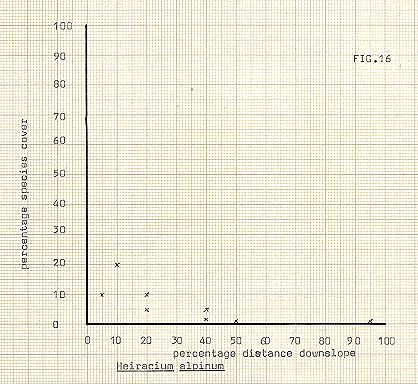
Figure 16
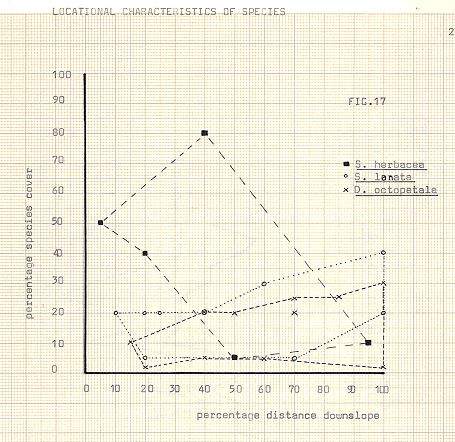
Figure 17
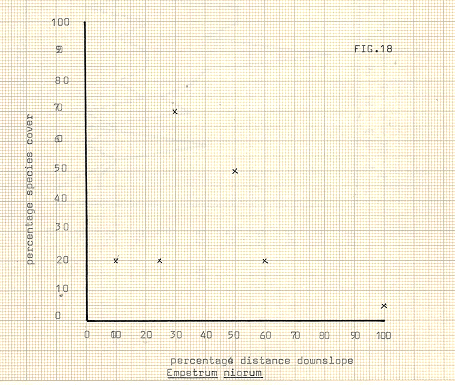
Figure 18
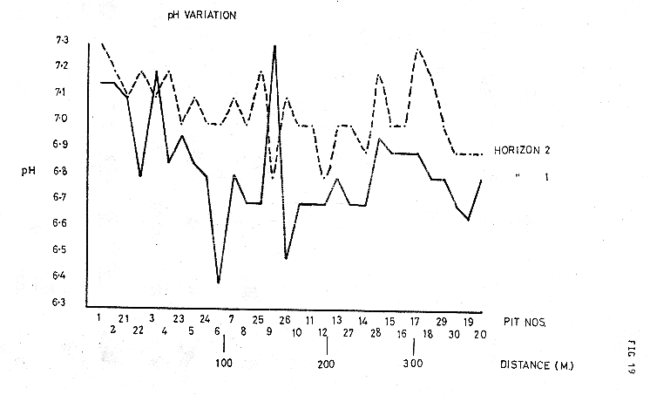
Figure 19
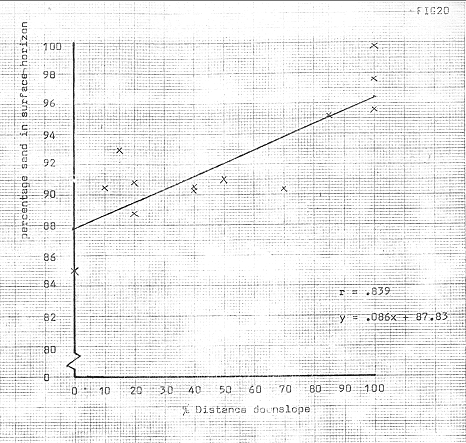
Figure 20
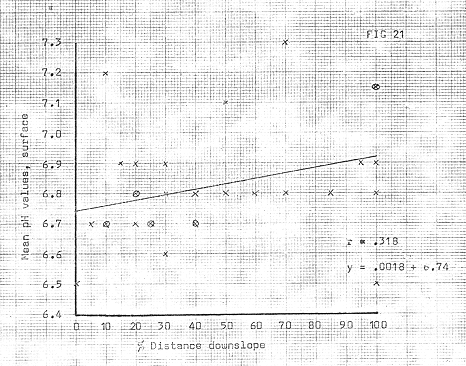
Figure 21
Figure 22
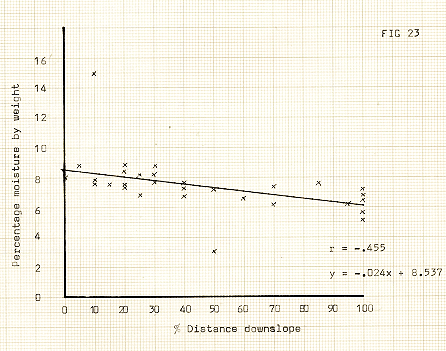
Figure 23
Table 2
Table 3
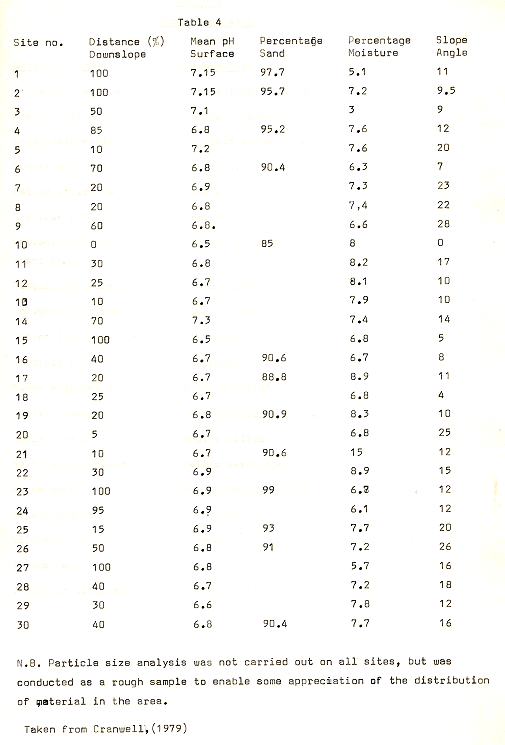
Table 4
Table 5
Characteristics of plant species
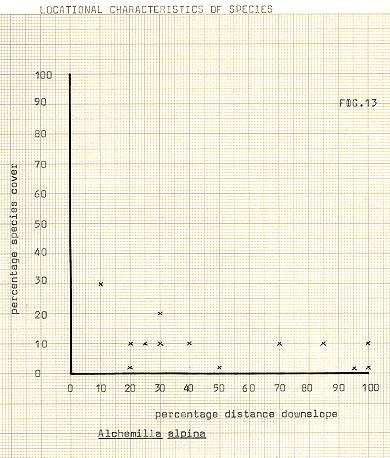
Alchamilla alpina – perennial; inhabitant of dry slopes in naked places, common in Iceland
Arabis alpina – perennial; inhabitant of damp places, often with mass and on small rivulets; common over most of Iceland
Dryas octopetala – perennial; occurs upon heaths, and in gravelly places; very common in Iceland
Empetrum nigrum – perennial; occurs on heaths, moors etc.; common in Iceland
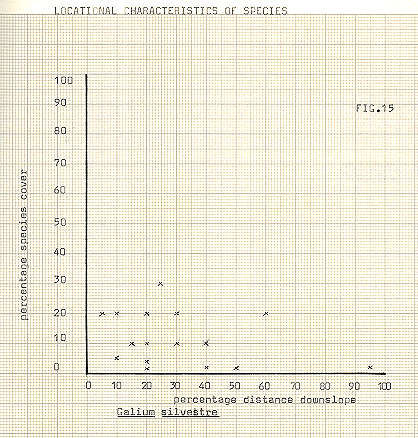
Galium silvestre – perennial; occurs on dry, sandy grasslands, lava fields very common in Iceland
Heiracium alpinum – perennial; occurs in grassy places, mountain slopes also sometimes on rocks; common in the hills in Iceland
Pinquicula vulqaris – perennial; occurs in damp of peaty soils, both on low and high ground, likes acidic conditions; common in all parts of Iceland
Polygonum viviparum – perennial; occurs on pastures, heaths, and on slopes very common in Iceland
Potentilla rupestris – perennial; occurs on sandy, sometimes turfy soil near to the sea; common in Iceland
Rhodiola rosea – perennial; occurs in rock fissures, open gravelly soils small size on open ground; common in Iceland
Rumex acetosella – perennial; occurs on open, sandy ground; common in the South and South-West of Iceland
Salix herbacea – perennial; occurs on gravelly ground; common throughout Iceland
Salix lanata – perennial; occurs on dry, open ground, where it often dominates; not rare in Iceland
Saxifraga aizoides – perennial; occurs commonly in wet places; rather common in Eastern IcelandSedum acre – occurs in gravelly, rather dry places; rather common in Iceland
Silene acaulis – perennial; very common oh rock ledges, open ground, often right down to the sea; very common in Iceland
Silene vulgaris – perennial; occurs as a weed, introduced near villages very rare in Iceland
Thymus serpyllum – perennial; occurs on hillslopes, and on dry, sandy or gravelly ground; common in Iceland
Vaccinium uliginosum – perennial; native of bogs and heather moors, also common on hillslopes; common in Iceland
(Largely from Ostenfeld and Grontved; 1934)
Discussion
As was noted in the introduction, the relief of the area seemed to play, and was later demonstrated to play, a vital role in the soil forming processes of the area, (Cranwell, 1979). It seemed likely to follow then, that the pattern of vegetation of the area would be equally strongly affected by the complexity of the terrain, and it comes as no great surprise that the “borders” of the communities were to a large extent determined by the same factors. The communities which were identified by the association analysis were generally the ridge colonisers, and the hollow colonisers, with gradations on the slopes in between. The results that are presented in the preceding section therefore lead us to consider the microenvironments of these communities.
Micro-environments
Ridge tops
The summits of the ridges were characterised by relatively high concentrations of silt size particles, and some indication of this feature can be seen from figure 20. This was primarily due to a cumulative causation process, involving the initial exposure of surfaces to low winter temperatures, which tended to lead to frost heaving. Krynine and Judd, (l957) assert that concentrations of over 3% of silt and clay sized particles leads to frost heaving in this type of environment, and given that some sites showed concentrations of up to 15%, frost heaving would be likely to be spectacular.
This in turn leads to a movement of stones and larger particles upward through the profile to the surface, where, due to the continuing disturbance from below, causes larger particles to gravitate downslope into the hollows. Once the pattern is established, the process is self-reinforcing, primarily due to the higher capacity of silt-sized particles for holding water by adsorption to their surfaces, which makes them more susceptible to the growth of ice crystals and lenses between the particles, termed ice segregation (for a fuller explanation, see Cooke and Doornkamp, (1974). The growth of the crystals themselves generates high internal pressures between particles and precipitates heaving thus, the characteristics of ridge tops are dominated by frost-heave.
A further point to note is that the higher concentrations of silt sized particles causes higher (relative) moisture contents in the ridge tops, although the general pattern is for a low moisture content c 5-9% by weight. The concentration also leads to relatively higher bulk density values on the ridge tops. 0n occasions, this density exceeded 2000 kg/m3, which according to some authors is the upper limit for plant growth, (Donahue, Shickluna & Robertson, 1971). The primary reason for this cycle is the blow-out of snow cover from the ridges, thus removing any insulation from the freezing winter temperatures.
As can be expected, these environments were the least abundant in vegetation, and most of the species found in the transect as a whole were absent from these harsh areas. Predominant cover was from the shrubby perennials, but some herbaceous perennials tended to become more evident as the soils stabilised and became more insulated under vegetation on a gradient away from the glacier snout.
The hollows
Hollows in the moraine topography were noticeably higher both in vegetation cover and in number of species present, as can be seen from figure 6. The hollows of course were much more protected habitats, being insulated from freezing winter temperatures by the snow cover which accumulated there, in contrast to the exposed ridges It seems likely that desiccation, too, was suffered more severely by the ridge colonisers, since the hollows tended to reduce windspeeds although the hollows may have increased turbulence somewhat.
The pH values were generally higher than the ridge tops, though usually less than .5 of a unit (see fig 21), despite the effects of organic acids produced by the vegetation; this seems primarily due to the effects of leaching on the bare upper slopes, a feature which has previously been noted by Crocker, (1960), in the Glacier Bay sequences where decrease of pH by leaching often exceeds the rate of decrease under vegetation. In conjunction with this, a general decrease of pH was noted, with distance from the ice front; (see fig 19).
In some hollows an accumulation of larger stones had occurred, in concurrence with features noted by Tisdale et al; (l966), which they identified as solution hollows. Channels on be surface of the glacier when it was at that level of advance tended to accumulate debris from the surface and, as the glacier thinned at the snout, the debris was dumped. However, these tended to be few in number, and the general pattern was for a mixture of different sized particles to be evident in the hollows, not least due to the high rates of inwashing of materials from the slopes, with high runoff rates, severe gullying and in some cases, solifluction.
The slopes
Slope angles varied between 0 and 35 degrees in the area, with a predominance of higher angled slopes especially near the snout, however, the slope angles at the sites themselves were more in the range of 5-20 degrees, (see figure 22). This, in combination with the unconsolidated nature of the materials, and what appeared from purely cursory knowledge to be an area of high rainfall, meant high erosion rates on the surfaces of slopes, especially. Extensive gullying was clearly evident, some over a metre deep. In periods of heavy or extended rainfall, spontaneous flows of material were observed. Vegetation on the steeper slopes probably inhabited the most insecure of habitats, if only in the previously mentioned respects, however, in addition, the sporadic dislodgement of stones from upslope presented a threat to any but the most low-lying of species.
In general, the habitats offered by the moraines was a singularly inhospitable one, in terms of climate, slope instability and length of growing season, however a further problem of existence, that of nutrient supply was of importance, I suspect. Although no tests were made, previous work by Crocker and Major, (1955), has shown that some nutrients, and especially Nitrogen are very limiting, despite a general availability of exchangeable rations, and it is the short supply of this essential element which principally causes the slow rate of colonisation on raw tills.
The Communities
Communities 1- 3; figures 7- 9
The ridge colonisers, as I have chosen to call them, occupy a very unstable habitat, coupled with environmental extremes. New individuals face considerable difficulty in becoming established here, but the prostrate shrubs seem to have an advantage, once established. Their origin may have been in droppings on ridge top perches, since the seeds of V.uliginosum and E.nigrum are often food targets for birds. One advantage of the prostrate shrub in this environment is that of permanence. The yearly maintenance costs in carbohydrates are low, and reproduction need only occur at long intervals, (Ives and Barry; 1974). Thus, the long lived shrub provides some flexibility in meeting the short-term changes in the environment. Hardley and Bliss, (1964), have also suggested that older evergreen leaves may act as winter storage organs, since lipids and proteins are mobilised and their stored energy translocated to new leaves in the growing season. The mats of the shrubs help to stabilise the ridge-tops to some degree, once established, and it is at this stage that later colonisers appear, S.acre and R .rosea. although of rare occurrence in this environment, seem to form a distinct association with E. nigrum, although this may possibly be due to their limited distribution, (viz, 1 each ), than to any distinct communality, despite the fact that they occur together.
Arabis alpina seems to form a more sound association with mosses, (community no 2, fig 8 ), occurring high on the least exposed slopes and in relatively sheltered positions. Spores of the mosses are obviously fairly ubiquitous, partly due to their commonness in the surrounding area, but establishment in large patches only occurs where they are not in direct competition with other mote vigourous plants. Because of their carpeting strategy, mosses can survive the disturbances of rocks falling downslope, and can also survive all but the most damaging of gullying, and on slopes the mosses are assured of a good supply of moisture and of nutrients being washed downslope.
Arabis alpina, too is a low plant, forming mats with extensive runners and in this way is well adapted to a potentially disturbed environment. It seems likely that strong competition from the more numenous species of the hollows is too great, by the time this community is established, which is generally at later stages on the moraines. The communities of higher slopes show quite characteristic traits in many cases. The high rainfall in the area does not prevent water shortages, which result from the rapid drainage in the granular and unconsolidated material of the moraines, thus the woody and extensive rooting systems of the Ericaceae present proves a successful strategy. Alternatively, the succulent type strategy of species such as S.acre and R.rosea may overcome periods of dryness without damage by conservation and storage. The mosses themselves are also able to withstand long periods of drought by an enforced dormancy, being rapidly rejuvenated on the advent of precipitation. The relative paucity of species on the higher slopes contrasts strongly to greater or lesser degree with the diversity of the hollows. The majority of sites where many species were present showed low cover rates per species, although in a few hollows some competitive advantage was being exploited well, with one or two species becoming dominant, with other species being present only incidentally.
Dryas and Salix usually occupied the protagonist’s role in these situations, although in two instances a singularly dominant cover of mosses was recorded, (see Table 1). The sites where many species tended to occur, (community no 4, fig 10), in the older moraines often contained species from the higher slopes too, which partially explains the wide scatter of points in figure 10. However, as has been noted, these species often formed communities of their own, and this is an example of the breadth of their niches. A major factor in explaining their presence is undoubtedly the increased availability of nutrients from the inwashing of material from upslope. However, extension of niche breadth seems to have occurred in an upslope direction for these species, rather than the reverse. This particular community comes under scrutiny later in the discussion in the section on succession.
The remaining two communities, numbers 5 and 6, seem to be relatively marginal, community no 5, (figure 11), consists of Saxifraga aizoides and P. rupestris, both of which are widely scattered, but with a preponderance towards the higher slopes, where they both appear as isolated groups of individuals with low percentage cover. Both these species probably suffer in competition with the more vigourous species occupying the hollows, which may explain their predilection for the rocky, upper slopes.
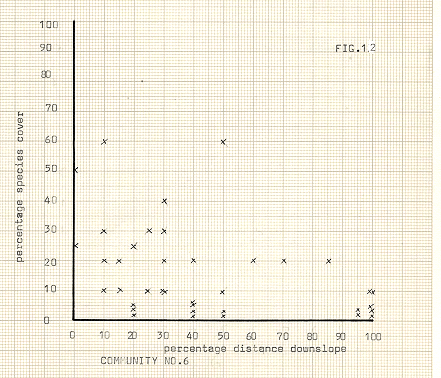
Community number 6, (figure 12), is made up of Graminae, Silene acaulis and T.serpyllum, There is clearly a bias toward the upper slopes here too, although both the spread of individuals, and the degree of cover is higher than in community no.5. T.serpyllum often occurs in isolated dense mats, in much the same strategy as Musci spp., and achieves a high degree of cover, as does Silene, although to a lesser extent. The selection by the program of the distinction between the communities 5 and 6 is a difficult one to reconcile with observations, since so many similarities in slope location and range of cover on slopes exist, it is almost impossible to envisage a valid reason for explaining this distinction.
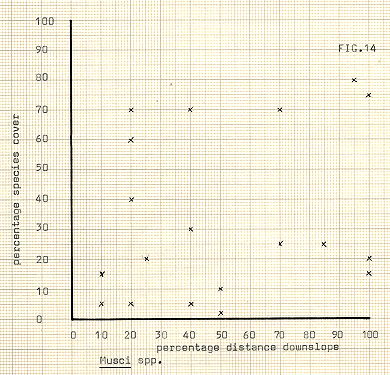
In attempting to delineate differences between the communities, the various species upon which division took place have been illustrated in figures 13-18, against distance downslope. In some cases, notably Galium, Heiracium and Empetrum, a quite noticeable bias towards a preferred slope position occurs. Bn the other hand, species such as the mosses seem to show a very variable occurrence, and cover, in relation to distance downslope. The fact that individual species may not display strong characteristic in location does not of course necessarily infer that the communities previously referred to are any less valid, since the most important feature of the community analysis is that of mutual association. In any event it appears to be the usual case for divisions to occur first upon species which are the most, or among the most, widespread in an area.
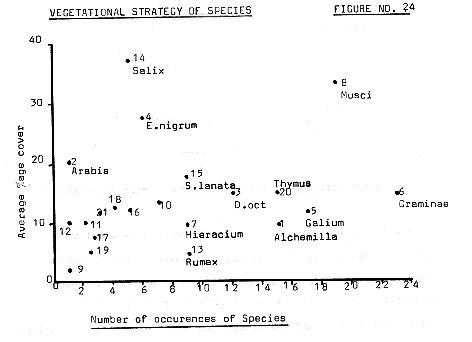
One interesting aspect of the vegetation of the area is illustrated in figure 24, overleaf, showing the percentage cover against the number of occurrences of the various species. As can be seen quite clearly, low cover rates are recorded for the majority of species, but some, like Salix, (the two varieties have been put separately to clarify their strategies ), have quite differing characteristics even within the same species. S.herbacea occurs quite infrequently, but where it occurs, it appears to dominate the vegetation, having a high average cover rate, whereas S.lanata tends to occur more frequently, whilst having lower cover rates generally. In fact, as shall be shown in the section on succession, the two species occupy quite different roles
Figure 24
Number of occurrences of Species
The species which occur most frequently are perhaps the species which posses the best dispersal mechanisms; mosses for instance, are at a considerable advantage when one considers the lightness and mobility of their spores. In the same way the grasses, which were thought to be of Festuca genus, can be quite mobile, along with the feathered seeds of Dryas. The concentration of species having both low occurrences and cover rates seems to reflect in part at least, the situation in the hollows, previously mentioned, where a larger number of species with low cover rates are competing in the hollows.
A further aspect of some interest is the form of the plants in such a harsh environment. Some species, such as Thymus. tend to prefer the prostrate, mat-forming strategy for stability and access to water in the rapidly draining soils, whilst others, such as Polygonum, may prefer the deep rooting strategy.
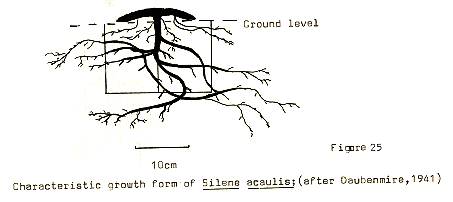
Figure 25
In any event, all of the species found tend to have the majority of their mass underground, as protection from the more severe conditions which he aerial parts have to endure. In this respect, it seems quite clear that species having similar growth forms are most likely to induce competition between themselves, and in view of the lack of diversity in the environment, it will probably mean extinction for one species or the other, since narrowing of niche breadth is hardly possible in such a stressed situation. On the other hand, where a number of species exist, having differing growth forms, then less interference is likely to occur. It is in this way perhaps that such a relatively large number of species may be found in small pockets of the hollows, where a number of individuals with growth forms such as that of Polygonum may coexist alongside mat forming species such as Dryas. In fact it is this very notion of coexistence or competition which leads us on to consider the question of succession.
Figure 26
Succession
From the works of Cooper, (1916), and Tisdale, (1966), which seem to be the most specific works on the nature of succession behind retreating glaciers, one major point may be drawn. They regard the successional stages as being characterised by the Dryas and the Salix stages, followed by a gradual approach to the composition of the surrounding forest. In the Svinafell moraines it prove to be not quite as clear cut as one might have supposed from reading the works mentioned above, however, similar trends did in fact exist, although in fact, the main scene for the battle of succession occurred in the hollows, and relatively few changes were observed on the higher slopes, although, as has been noted, the amelioration of surface conditions on the ridges and higher slopes by species such as Empetrum did in fact allow colonisation by other species which were previously unable to survive the difficult conditions on the ridges.
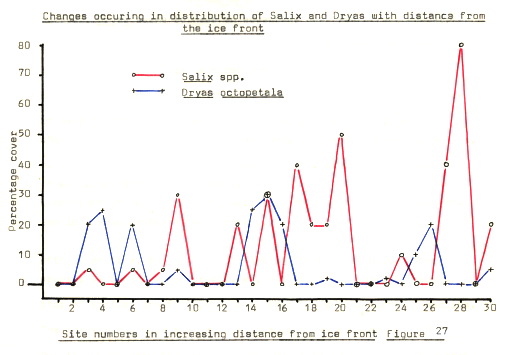
Figure 27
In figure 27. above, a general pattern of increasing cover of the Salix species can be observed, after an initial, though less detectable dominance by Dryas. Dryas itself has a considerable advantage over most of the other species found in the area since, in an area where Nitrogen is usually the most limiting factor, the presence of Nitrogen fixing bacterial nodules on the roots must be of considerable importance in establishing, and maintaining plants in this area. Lawrence, (1958), Crocker, (1960), and Tisdale, (1966), have all mentioned the presence of these nodules on Dryas, and Crocker, (1960) considered the nodules to be of primary importance in improving the Nitrogen status of these soils.
The existence of these nodules must be important whilst the nutrient status of the soils is very low, once the plant is established, however, Dryas seems to have an advantage over its direct competitors in the establishment phase too. The multitude of feathered, light seeds which Dryas produces are capable of widespread dispersion over the area, and, with such a potential, it seems reasonable to expect that Dryas should be widespread. However, as has been shown, Dryas is not the most widespread, at least amongst the sample, but it does occur in virtually every hollow, since the windspeeds on be ridges do not allow the light seeds to fall, but the stiller conditions of the hollows may well do, and thus encourage propagation in a more protected environment.
After some time, the soil conditions in the hollows allow other species to become more established, but it is not until a competitor of similar growth form appears that Dryas show signs of decline. The competitor, from available information was assumed to be Salix, but it now appears that only one of the Salix varieties is in direct competition with Dryas. and that is S.lanata. Particularly of use in illustrating this is the graph showing locational characteristics in figure 17, where S.herbacea quite clearly occupies a different area in terms of distance downslope, than does S.lanata, whose sphere of influence almost exactly coincides with that of Dryas itself.
It seems therefore, from available evidence, then, that Dryas is a comparatively successful “R” strategist, in relation to the Salix species. Their nitrogen-fixing bacterial nodules apparently give them a competitive advantage, even though Salix is present from even the earliest stages of colonisation. Salix lanata, the main direct competitor, appears to be a slower growing plant but its competitive ability increases with time, as the advantage of Dryas becomes of less importance. It is possible that the greater vigour of S.lanata, which eventually achieves expression, is due to some hybrid vigour resulting from combination with S.herbacea, however, this ability is delayed somewhat by the extreme shortage of nutrients in the environment, and to some extent is masked by the much greater initial vigour of Dryas.
Conclusion
It seems apparent that the use of association analysis has been of advantage in identifying the various communities of the. area, but this use has been subject to some problems caused by the influence of two major variables in the sample.
Firstly, the variability of habitat, (and therefore, largely of communities) with the topography of the area; and secondly, the Variation of the vegetation with time, or, in this case distance which has largely been seen as a surrogate for time, have both had to be accommodated in be analysis.
For the most part, the analysis itself has not brought out the effect of the second variable, whilst it has quite effectively handled the first, and major, determinant of the vegetation of the area. It has been demonstrated that significant variations in micro-environment exist within the area, and that as a result, particular communities develop which are peculiar to those microenvironments.
However, the subject of succession, since it is an area which is effectively time-based, has been to a great extent ineffectively explored by the association analysis, almost certainly due to the overriding influence of the topography, therefore treatment of the subject has largely been of the nature of inference from the original data. This particular line of enquiry has been concerned mainly with examining the validity of those stages of succession suggested by other workers in the field.
In this respect, the succession of Dryas by Salix has been narrowed down to the identification of the particular Salix species which is of most influence in the successions, that of S.lanata. Some consideration too, of the way in which this succession takes place has been given, especially in regard to the location of the successions and the strategies of the species involved which give rise to the succession. It is clear that other changes occur which at present are beyond the scope of this work, changes of a successional nature occurring in other locations within the moraines. It is customary to allude to the potential for other work in the area covered and in this respect, I have two major suggestions, both in their ways arising from the inadequacies of this study.
Firstly, in order to better delineate the characteristics of a particular community, it seems superfluous to use some common method of identifying characteristics of a spread of points, such as a regression line, and in fact this is why I have not done this. Much more of use would be some form of numerical analysis leading to a fitting of a curve which is peculiar only to that community, in the way that, for example, electrocardiograms may be characteristic of some particular heart ‘disease or condition. Secondly, if any more detailed data are required upon this area of work, it seems all too obvious that a much larger data base is required, although for the purposes of this study the data has proven to be of adequate proportions in terms of bulk.
As a final note, for would-be students of successions in any situation, the study of glacial sequences has a great deal to offer, much of which has not been used effectively in the past and it is to be hoped that such omission will be less frequent in the future.
Bibliography
Cooke, R .U. , & Doornkamp, J.C. (1974), Geomorphology in environmental management; O.U.P.